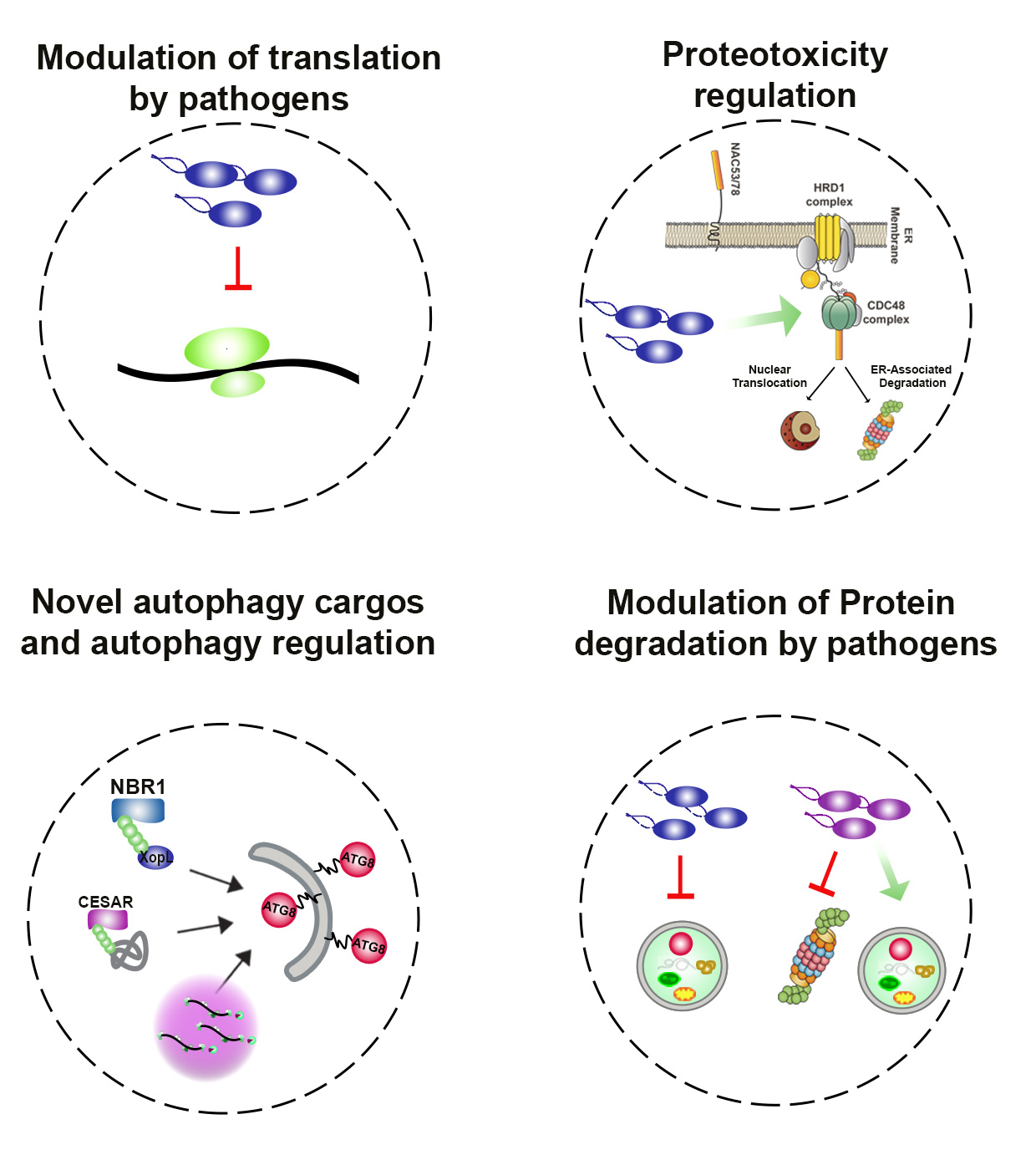
 Our lab’s recent key finding revealed novel mechanisms and components of proteostasis, as well as new strategies of microbes to perturb plant proteostasis: we show for the how bacterial effectors undergo phase-separation to target processing body (PB) – dependent translational arrest. We also reveal evidence that uncovers a previously unknown role of autophagy in regulating PBs and influencing protein translation (González-Fuente et al., 2025). Thus, our recent discoveries demonstrate that pathogens manipulate proteostasis at every step, from protein translation (González-Fuente et al., 2025), trafficking (Brinkmann et al., 2024) to protein degradation (Leong et al., 2022).
Our lab’s recent key finding revealed novel mechanisms and components of proteostasis, as well as new strategies of microbes to perturb plant proteostasis: we show for the how bacterial effectors undergo phase-separation to target processing body (PB) – dependent translational arrest. We also reveal evidence that uncovers a previously unknown role of autophagy in regulating PBs and influencing protein translation (González-Fuente et al., 2025). Thus, our recent discoveries demonstrate that pathogens manipulate proteostasis at every step, from protein translation (González-Fuente et al., 2025), trafficking (Brinkmann et al., 2024) to protein degradation (Leong et al., 2022).
We have identified a novel selective autophagy pathway, effectorphagy, a form of animal xenophagy, where microbial effectors are selectively degraded by autophagy to dampen infection. However, pathogens can revert this process by suppressing degradation machineries (Leong et al., 2022; Üstün et al., 2018). As such, manipulation of proteostasis by pathogens induce proteotoxic stress, the accumulation of abnormal proteins, which can cause irreversible cellular damage. However, little was known how proteotoxicity caused by different sources is regulated in eukaryotes. Our recent findings in Langin et al., 2024 BioRxiv highlight that proteasomal transcriptional activators NAC53/78 co-ordinate the response to proteotoxicity via ER-anchored protein sorting (ERAPS). Our study uncovers that both transcription factors orchestrate the communication between the nucleus and chloroplast during various stress conditions. To date no similar direct mechanism has been deciphered linking the proteasome and energetic metabolism in other systems.
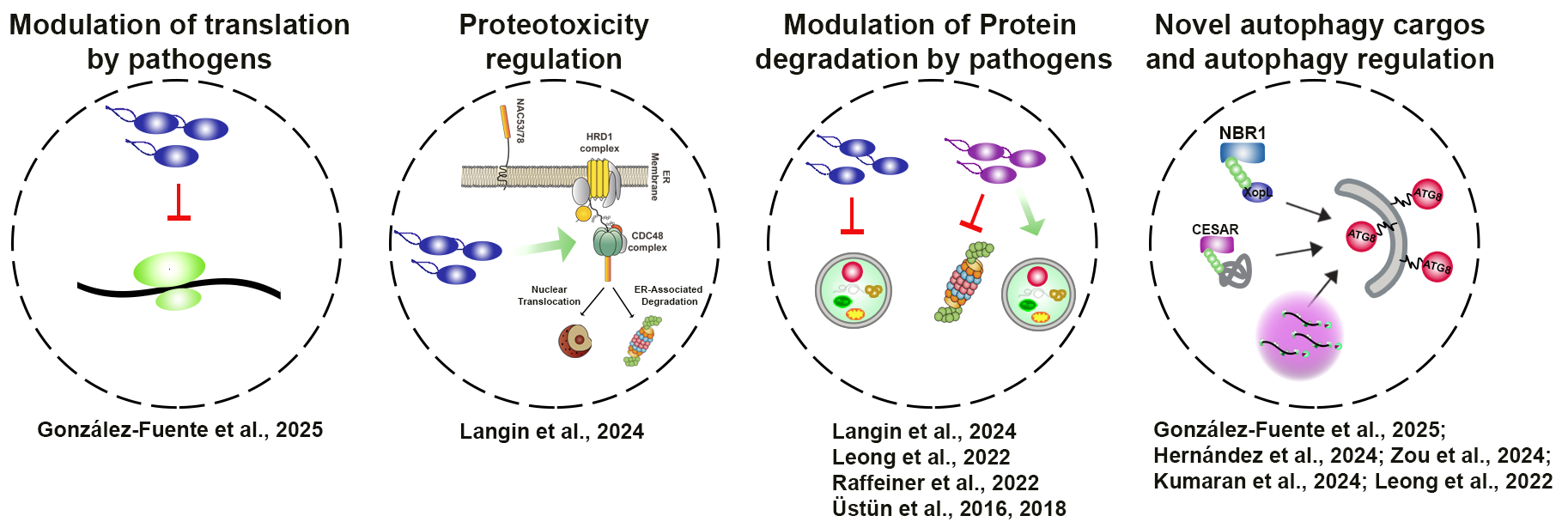
We have made other important contributions in how proteotoxicity via autophagy is regulated in Arabidopsis, tomato and Chlamydomonas (Zou et al., 2025; Hernández et al., 2025; Kumaran et al., 2025). Taken together, we have established ourselves in the field of proteostasis research focusing on pathogens that manipulate proteostasis at every step to cause proteotoxicity and also identified novel proteostasis cargos and regulators.
